Классификация мутаций по изменениям в кодирующей последовательности ДНК (замены, вставки, делеции). Изменение собственных генов всего лишь одной инъекциeй – если, конечно, вы везучи Изменение набора генов
Мутации (от лат. mutatio - изменять) - это передава емые по наследству структурные изменения генов.
Крупные мутации (геномные перестройки) сопро вождаются выпадением или изменением относительно крупных участков генома - такие мутации, как правило, необратимы.
Мелкие (точковые) мутации связаны с выпадением или добавлением отдельных нуклеотидов ДНК. При этом изменяется лишь небольшое число признаков. Такие измененные бактерии могут полностью возвращаться в исходное состояние (ревертировать).
Бактерии с измененными признаками называются му тантами. Факторы, вызывающие образование мутантов, носят название мутагенов.
Бактериальные мутации делят на спонтанные и индуци рованные. Спонтанные (самопроизвольные) мутации возникают под влиянием неконтролируемых факторов, т. е. без вмешательства экспериментатора. Индуциро ванные (направленные) мутации появляются в результа те обработки микроорганизмов специальными мутагенами (химическими веществами, излучением, температурой и
В результате бактериальных мутаций могут отмечаться: а) изменение морфологических свойств б) изменение культуральных свойств в) возникновение у микроорганизмов устойчивости к лекарственным препаратам г) потеря способности синтезировать аминокислоты, утилизировать углеводы и другие питательные вещества д) ослабление болезнетворных свойств и т. д.
Если мутация приводит к тому, что мутагенные клетки обретают по сравнению с остальными клетками популяций преимущества, то формируется популяция из мутантных клеток и все приобретенные свойства передаются по наследству. Если же мутация не дает клетке преимуществ, то мутантные клетки, как правило, погибают.
Трансформация. Клет ки, которые способны воспринять ДНК другой клетки в процессе трансформации, называются компетентными.
Трансдукция - это перенос генетической информа ции (ДНК) от бактерии донора к бактерии реципиенту при участии бактериофага. Трансдуцирующими свойствами обладают в основном умеренные фаги. Размножаясь в бактериальной клетке, фаги включают в состав своей ДНК часть бактериальной ДНК и передают ее реципиенту.
Различают три типа трансдукции: общую, специфи ческую и абортивную.
1 . Общая трансдукция - это передача различных генов, локализованных на разных участках бактериальной хромосомы.
При этом бактерии доноры могут передать реципиенту разнообразные признаки и свойства- способность образовывать новые ферменты, устойчивость к лекарственным препаратам и т. д.
2. Специфическая трансдукция - это передача фагом только некоторых специфических генов, локализованных на специальных участках бактериальной хромосо мы. В этом случае передаются только определенные признаки и свойства.
3. Абортивная трансдукция - перенос фагом ка кого-то одного фрагмента хромосомы донора. Обычно этот фрагмент не включается в хромосому клетки реципиента, а циркулирует в цитоплазме. При делении клетки реципиента этот фрагмент передается только одной из двух дочерних клеток, а второй клетке достается неизме ненная хромосома реципиента.
С помощью трансдуцирующих фагов можно передать от одной клетки другой целый ряд свойств, таких как способность образовывать токсин, споры, жгутики, проду цировать дополнительные ферменты, устойчивость к лекарственным препаратам и т. д.
Конъюгация - это передача генетического матери ала от одной бактерии к другой при непосредственном контакте клеток. Клетки, передающие генетический мате риал, называются донорами, воспринимающие его - реципиентами. Этот процесс носит односторонний характер - от клетки донора к клетке реципиента.
Бактерии донора обозначаются F + (мужской тип), а бактерии реципиента - F - (женский тип). При тесном сближении клеток F + и F - между ними возникает цитоплазматический мостик. Образование мостика контро лируется фактором F (от англ. Fertility - плодовитость). Этот фактор содержит гены, ответственные за образова ние половых ворсинок (sex - pili). Функцию донора могут выполнять только те клетки, которые содержат фактор F . Клетки реципиента лишены этого фактора. При скрещива нии фактор F передается клеткой донора реципиенту. Получив фактор F , женская клетка сама становится донором (F +).
Процесс конъюгации можно прервать механическим способом, например встряхиванием. В этом случае реципи ент получает неполную информацию, заключенную в ДНК.
Конъюгация, как и другие виды рекомбинации, может осуществляться не только между бактериями одного и того же вида, но и между бактериями разных видов. В этих случаях рекомбинация называется межви довой.
Плаз миды - это сравнительно небольшие внехромо-сомные молекулы ДНК бактериальной клетки. Они расположены в цитоплазме и имеют кольцевую структуру. В плазмидах содержится несколько генов, функциониру ющих независимо от генов, содержащихся в хромосомной ДНК.
Профаги, вызывающие у лизогенной клетки ряд изме нений, передающихся по наследству, например способ ность образовывать токсин (см. трансдукцию).
F -фактор, находящийся в автономном состоянии и принимающий участие в процессе конъюгации (см. конъ югацию).
R -фактор, придающий клетке устойчивость к лекар ственным препаратам (впервые R -фактор был выделен из кишечной палочки, затем из шигелл). Исследования показали, что R -фактор может быть удален из клетки, что вообще характерно для плазмид.
К-фактор обладает внутривидовой, межвидовой и даже межродовой трансмиссивностью, что может явиться причиной формирования трудно диагностируемых атипичных штаммов.
Бактериоциногенные факторы (col -факторы), которые впервые были обнаружены в культуре кишечной палочки (E . coli), в связи с чем названы колицинами. В дальней шем они были выявлены и у других бактерий: холерного вибриона - вибриоцины, стафилококков - стафилоцины и др.
Со l -фактор - это маленькая автономная плазмида, которая детерминирует синтез белковых веществ, способ ных вызывать гибель бактерий собственного вида или близкородственного. Бактериоцины адсорбируются на поверхности чувствительных клеток и вызывают нарушения метаболизма, что приводит клетку к гибели.
В естественных условиях только единичные клетки в популяции (1 на 1000) спонтанно продуцируют колицин. Однако при некоторых воздействиях на культуру (обработка бактерий УФ-лучами) количество колицинпродуцйрующих клеток увеличивается.
ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ ИЗМЕНЧИВОСТИ микроорганизмов
Еще Пастер искусственным путем получил необратимые изменения у возбудителей бешенства, сибирской язвы и приготовил вакцины, предохраняющие от этих заболеваний. В дальнейшем исследования в области генетики и изменчивости микроорганизмов позволили получить большое число бактериальных и вирусных штаммов, используемых для получения вакцин.
Результаты исследования генетики микроорганизмов с успехом были использованы для выяснения закономерностей наследственности высших организмов.
Большое научное и практическое значение имеет также новый раздел генетики - генная инженерия.
Методы генной инженерии позволяют изменять структуру генов и включать в хромосому бактерий гены других организмов, ответственных за синтез важных и нужных веществ. В результате микроорганизмы становятся продуцентами таких веществ, получение которых химическим путем представляет очень сложную, а иногда даже невозможную задачу. Этим путем в настоящее время получают такие медицинские препараты, как инсулин, интерферон и др. При использовании мутагенных факторов и селекции были получены мутанты-продуценты антибиотиков, которые в 100-1000 раз активнее исходных.
9. Генетика иммунитета
Генетическая детерминированность иммунной реакции организма высших животных
Механизм синтеза моноспецифических антител и иммунная память
Наследуемость уровня иммунной реакции организма и возможности селекции животных по устойчивости к инфекциям.
Иммунитет – это невосприимчивость организма к инфекционным агентам и генетически чужеродным веществам антигенной природы. Главная функция иммунитета – иммунологический надзор за внутренним постоянством (гомеостазом) организма.
Следствием этой функции является распознавание, а потом блокирование, нейтрализация или уничтожение генетически чужеродных веществ (вирусов, бактерий, раковых клеток и т.д.). За сохранение генетически обусловленной биологической индивидуальности отвечает иммунная система организма – совокупность всех лимфоидных клеток (специфический фактор защиты). К неспецифическим факторам защиты относят кожные и слизистые покровы. Иммунный ответ, или иммунологическая реактивность – форма реакций организма на чужеродные вещества (антигены). Главной функцией антител является их способность вступать в быструю реакцию с антигеном в виде реакции глютинации, преципитации, лизиса, нейтрализации.
10. Группы крови и биохимический полиморфизм.
Понятие о группах крови
Наследуемость групп крови
Практическое применение групп крови в животноводстве
Полиморфные системы белков и их связь с продуктивностью животных
Методы определения групп крови и полиморфных систем белков.
Группы крови были открыты в 1900 г. (у человека) и объяснены в 1924 г. А в 1936 году использован термин иммуногенетика. В пределах вида особи различаются по ряду охимических, генетически детерминируемых признаков, которые могут быть выявлены иммуногенетически в виде антигенов (генетически чужеродные вещества, при введении их в организм вызывают иммуногенетических реакций). Антитела – иммуноглобулины (белки), образующие в организме под воздействием антигенов, различия в групповой принадлежности крови определяются антигенами, расположенными на поверхности эритроцитов. Антигенные факторы иногда называют кровяными факторами, сумму всех групп крови одной особи – типом крови. После рождения группы крови у животных не меняется. Генетические системы групп крови и антигены обозначают прописными и строчными буквами – А,В,С и т.д. Количество антигенов много, поэтому пишут со значками А, В, С, и с подстрочными индексами А1, А2 и т.д.
Главная | О нас | Обратная связь
ГЕНОТИПИЧЕСКАЯ (НАСЛЕДУЕМАЯ) ИЗМЕНЧИВОСТЬ
Генотипическая изменчивость может возникать в результате мутаций и генетических рекомбинаций.
Мутации (от лат. mutatio - изменять) - это передаваемые по наследству структурные изменения генов.
Крупные мутации (геномные перестройки) сопровождаются выпадением или изменением относительно крупных участков генома -такие мутации, как правило, необратимы.
Мелкие (точковые) мутации связаны с выпадением или добавлением отдельных оснований ДНК. При этом изменяется лишь небольшое число признаков. Такие измененные бактерии могут полностью возвращаться в исходное состояние (ревертировать).
Бактерии с измененными признаками называются мутантами. Факторы, вызывающие образование мутантов, носят название мутагенов.
Бактериальные мутации делят на спонтанные и индуцированные. Спонтанные (самопроизвольные) мутации возникают под влиянием неконтролируемых факторов, т.е. без вмешательства экспериментатора. Индуцированные (направленные) мутации появляются в результате обработки микроорганизмов специальными мутагенами (химическими веществами, излучением, температурой и др.).
В результате бактериальных мутаций могут отмечаться:
а) изменение морфологических свойств
б) изменение культуральных свойств
в) возникновение у микроорганизмов устойчивости к лекарственным препаратам
г) потеря способности синтезировать аминокислоты, утилизировать углеводы и другие питательные вещества
д) ослабление болезнетворных свойств и т. д.
Если мутация приводит к тому, что мутагенные клетки обретают по сравнению с остальными клетками популяций преимущества, то формируется популяция из мутантных клеток, и все приобретенные свойства передаются по наследству. Если же мутация не дает клетке преимуществ, то мутантные клетки, как правило, погибают. Генетические рекомбинации. Трансформация. Клетки, которые способны воспринять ДНК другой клетки в процессе трансформации, называются компетентными. Состояние компетентности часто совпадает с логарифмической фазой роста.
Трансдукция - это перенос генетической информации от бактерии донора к бактерии реципиенту при участии бактериофага. Трансдуцирующими свойствами обладают, в основном, умеренные фаги. Размножаясь в бактериальной клетке, фаги включают в состав своей ДНК часть бактериальной ДНК и передают ее реципиенту. Различают три типа трансдукции: общую, специфическую и абортивную.
1. Общая трансдукция - это передача различных генов, локализованных на разных участках бактериальной хромосомы. При этом бактерии доноры могут передать реципиенту разнообразные признаки и свойства - способность образовывать новые ферменты, устойчивость к лекарственным препаратам и т. д.
2. Специфическая трансдукция - это передача
фагом только некоторых специфических генов, локализованных на специальных участках бактериальной хромосомы. В этом случае передаются только определенные признаки и свойства.
3. Абортивная трансдукция - перенос фагом какого-то одного фермента хромосомы донора. Обычно этот фрагмент не включается в хромосому клетки реципиента, а циркулирует в цитоплазме. При делении клетки реципиента этот фрагмент передается только одной из двух дочерних клеток, а второй клетке достается неизмененная хромосома реципиента.
С помощью трансдуцирующих фагов можно передать от одной клетки другой целый ряд свойств, таких как способность, образовывать токсин, споры, жгутики, продуцировать дополнительные ферменты, устойчивость к лекарственным препаратам и т. д.
Конъюгация - это передача генетического материла от одной бактерии к другой при непосредственном контакте клеток. Клетки, передающие генетический материал, называются донорами, воспринимающие его - реципиентами. Этот процесс носит односторонний характер - от клетки донора к клетке реципиента.
Бактерии донора обозначаются F+ (мужской тип), а бактерии реципиента - F- (женский тип). При тесном сближении клеток F+ и F- между ними возникает цитоплазматический мостик. Образование мостика контролируется фактором F (от англ. fertility -- плодовитость). Этот фактор содержит гены, ответственные за образование половых ворсинок (sex-pili). Функцию донора могут выполнять только те клетки, которые содержат фактор F. Клетки реципиента лишены этого фактора. При скрещивании фактор F передается клеткой донора реципиенту. Получив фактор F, женская клетка сама становится донором (F+).
Процесс конъюгации можно прервать механическим способом, например, встряхиванием. В этом случае реципиент получает неполную информацию, заключенную в ДНК.
Перенос генетической информации путем конъюгации лучше всего изучен у энтеробактерий.
Конъюгация, как и другие виды рекомбинации, может осуществляться не только между бактериями одного и того же вида, но и между бактериями разных видов, В этих случаях рекомбинация называется межвидовой.
Генотипическая изменчивость наследуемая
Плазмиды – это сравнительно небольшое внехромосомное молекулы ДНК бактериальной клетки. Они расположены в цитоплазме и имеют кольцевую структуру. В плазмидах содержится несколько генов, функционирующих независимо от генов, содержащихся в хромосомной ДНК.

Рис.54 Плазмиды (внехромосомные молекулы ДНК)
Типичным признаком плазмид служит их способность к самостоятельному воспроизведению (репликации).
Они могут также переходить из одной клетки в другую и включать в себя новые гены из окружающей среды. К числу плазмид относятся:
Профаги. вызывающие у лизогенной клетки ряд изменений, передающихся по наследству, например способность образовывать токсин (см. трансдукцию). F-фактор, находящийся в автономном состоянии и принимающий участие в процессе конъюгации (см. конъюгацию).
R-фактор, придающий клетке устойчивость к лекарственным препаратам (впервые R-фактор был выделен из кишечной палочки, затем из шигелл). Исследования показали, что R-фактор может быть удален из клетки, что вообще характерно для плазмид.
R-фактор обладает внутривидовой, межвидовой и даже межродовой трансмиссивностью, что может явиться причиной формирования трудно диагностируемых атипичных штаммов.
Бактериоциногенные факторы (col-факторы), которые впервые были обнаружены в культуре кишечной палочки (Е. coli), в связи с чем названы колицинами. В дальнейшем они были выявлены и у других бактерий: холерного вибриона - вибриоцины, стафилококков - стафилоцины и др.
Col-фактор - это маленькая автономная плазмида, которая детерминирует синтез белковых веществ, способных вызывать гибель бактерий собственного вида или близкородственного. Бактериоцины адсорбируются на поверхности чувствительных клеток и вызывают нарушения метаболизма, что приводит клетку к гибели.
В естественных условиях только единичные клетки в популяции (1 на 1000) спонтанно продуцируют колицин. Однако при некоторых воздействиях на культуру (обработка бактерий УФ-лучами) количество колицинпродуцирующих клеток увеличивается.
Изменения функциональных генов
По мутировавшим клеткам мутации могут быть соматические (например, разный цвет глаз у одного человека) и генеративные (или гаметические). Генеративные мутации передаются потомству, соматические проявляются у самой особи. Они передаются по наследству только при вегетативном размножении.
По исходу (значению) для организма выделяют мутации положительные, нейтральные и отрицательные. Положительные мутации появляются редко. Они повышают жизнеспособность организма и имеют значение для эволюции (например, мутации, приводящие к появлению четырехкамерного сердца в процессе эволюции хордовых). Нейтральные мутации практически не влияют на процессы жизнедеятельности (например, мутации, приводящие к наличию веснушек). Отрицательные мутации делят на полулетальные и летальные. Полулетальные мутации снижают жизнеспособность организма, сокращают срок его жизни (например, мутации, приводящие к болезни Дауна). Летальные мутации вызывают
смерть организма до рождения или в момент рождения (например, мутации, приводящие к отсутствие головного мозга).
По изменению фенотипа мутации бывают морфологические (например, уменьшенные глазные яблоки, шесть пальцев на руке) и биохимические (например, альбинизм, гемофилия).
По изменению генотипа выделяют мутации геномные, хромосомные и генные.
Геномные мутации – это изменение числа хромосом под действием факторов среды. Гаплоидия – набор хромосом 1n. В природе она встречается у трутней (самцов) пчел. Жизнеспособность таких организмов снижена, так как у них проявляются все рецессивные гены.
Полиплоидия – увеличение гаплоидного набора хромосом (3n, 4n, 5n). Полиплоидия используется в растениеводстве. Она приводит к повышению урожайности. Для человека гаплоидия и полиплоидия это летальные мутации.
Анеуплоидия – это изменение числа хромосом в отдельных парах (2n±1, 2n±2 и так далее).
Трисомия. например, если к паре половых хромосом женского организма добавляется Х-хромосома, развивается синдром трисомии Х (47, ХХХ), если она добавляется к половым хромосомам мужского организма, развивается синдром Клайнфельтера (47, ХХY). Моносомия. отсутствие одной хромосомы в паре – 45, Х0 – синдром Шерешевского-Тернера. Нулисомия. отсутствие пары гомологичных хромосом (для человека – летальная мутация).
Хромосомные мутации (или хромосомные аберрации) – это изменения структуры хромосом (межхромосомные или внутрихромосомные). Перестройки внутри одной хромосомы называются инверсии, нехватки (дефишенси и делеции), дупликации. Межхромосомные перестройки называются транслокации.
Примеры: делеция – синдром кошачьего крика у человека дупликация – появление полосковидных глаз у дрозофилы инверсия – изменение порядка расположения генов.
Транслокации могут быть: реципрокные – две хромосомы обмениваются сегментами нереципрокные – сегменты одной хромосомы переносятся на другую робертсоновские – две акроцентрические хромосомы соединяются своими центромерными участками.
Нехватки и дупликации всегда проявляются фенотипически, так как изменяется набор генов. Не всегда проявляются инверсии и транслокации. В этих случаях затрудняется конъюгация гомологичных хромосом и нарушается распределение генетического материала между дочерними клетками.
Генные мутации называются точковые, или трансгенации. Они связаны с изменениями структуры генов и вызывают развитие болезней обмена веществ (их частота 2-4%).
Изменения структурных генов.
1. Сдвиг рамки считывания происходит в случае выпадения или вставки одной или нескольких пар нуклеотидов в молекулу ДНК.
2. Транзиция – мутация, при которой происходит замена пуринового основания на пуриновое или пиримидинового на пиримидиновое (А Г или Ц Т). Такая замена приводит к изменению кодонов.
3. Трансверсия – замена пуринового основания на пиримидиновое или пиримидинового на пуриновое (А Ц Г Т) – приводит к изменению кодонов. Изменение смысла кодонов приводит к мисценс-мутациям. Если образуются бессмысленные кодоны (УАА, УАГ, УГА), они вызывают нонсенс-мутации. Эти кодоны не определяют аминокислоты, а являются терминаторами – они определяют конец считывания информации.
1. Изменен белок-репрессор, он не подходит к гену-оператору. В этом случае структурные гены не выключаются и работают постоянно.
2. Белок-репрессор плотно присоединяется к гену-оператору и не «снимается» индуктором. Структурные гены постоянно не работают.
3. Нарушение чередования процессов репрессии и индукции. Если индуктор отсутствует, специфический белок синтезируется, в присутствии индуктора он не синтезируется. Такие нарушения работы транскриптонов наблюдаются при мутациях гена-регулятора или гена-оператора.
В настоящее время описано около 5 000 болезней обмена веществ, причиной которых являются генные мутации. Примерами их могут быть фенилкетонурия, альбинизм, галактоземия, различные гемофилии, серповидно-клеточная анемия, ахондроплазия и др.
В большинстве случаев генные мутации проявляются фенотипически.
Наследственность и изменчивость. Хромосомная теория наследственности
Наследственность - это важнейшая особенность живых организмов, заключающаяся в способности передавать свойства и функции родителей потомкам. Эта передача осуществляется с помощью генов.
Ген - единица хранения, передачи и реализации наследственной информации. Ген представляет собой специфический участок молекулы ДНК, в структуре которого закодирована структура определенного полипептида (белка). Вероятно, многие участки ДНК не кодируют белки, а выполняют регулирующие функции. Во всяком случае в структуре генома человека только около 2% ДНК представляют собой последовательности, на основе которых идет синтез информационной РНК (процесс транскрипции), которая затем определяет последовательность аминокислот при синтезе белков (процесс трансляции). В настоящее время полагают, что в геноме человека имеется около 30 тыс. генов.
Гены расположены на хромосомах, которые находятся в ядрах клеток и представляют собой гигантские молекулы ДНК.
Хромосомная теория наследственности была сформулирована н 1902 г. Сэттоном и Бовери. Согласно этой теории хромосомы являются носителями генетической информации, определяющей наследственные свойства организма. У человека в каждой клетке имеется 46 хромосом, разделенных на 23 пары. Хромосомы, образующие пару, называются гомологичными.
Половые клетки (гаметы) образуются с помощью особого типа деления - мейоза. В результате мейоза в каждой половой клетке остается только по одной гомологичной хромосоме из каждой пары т.е. 23 хромосомы. Такой одинарный набор хромосом называется гаплоидным. При оплодотворении, когда сливаются мужская и женская половые клетки и образуется зигота, двойной набор, который называется диплоидным, восстанавливается. В зиготе у организма который из нее развивается, одна хромосома из каждой нары получена от отцовского организма, другая - от материнского.
Генотип - это совокупность генов, полученных организмом от родителей.
Другое явление, которое изучает генетика - изменчивость. Под изменчивостью понимают способность организмов приобретать новые признаки - различия в пределах вида. Выделяют две формы изменчивости:
- наследственную
- модификационную (ненаследственную).
Наследственная изменчивость - это форма изменчивости вызванная изменениями генотипа, которые могут быть связаны с мутационной либо комбинативной изменчивостью.
Мутационная изменчивость.
Гены время от времени подвергаются изменениям, которые получили название мутаций. Эти изменения имеют случайный характер и появляются спонтанно. Причины возникновения мутаций могут быть самыми разнообразными. Имеется целый ряд факторов воздействие которых повышает вероятность возникновения мутации. Это может быть воздействие определенных химических веществ радиации, температуры и т.д. С помощью этих средств можно вызывать мутации, однако случайный характер их возникновения сохраняется, и предсказать появление той или иной мутации невозможно.
Возникшие мутации передаются потомкам, т. е. определяют наследственную изменчивость, которая связанна с тем, где произошла мутация. Если мутация произошла в половой клетке то у нее есть возможность передаться потомкам, т.е. быть унаследованной. Если же мутация произошла в соматической клетке, то она передается только тем из них, которые возникают из этой соматической клетки. Такие мутации называются соматическими, они не передаются по наследству.
Различают несколько основных типов мутаций.
- Генные мутации, при которых изменения происходят на уровне отдельных генов, т. е. участков молекулы ДНК. Это может быть у трата нуклеотидов, замена одного основания на другое, перестановка нуклеотидов или добавление новых.
- Хромосомные мутации, связанные с нарушением структуры хромосом, приводят к серьезным изменениям, которые могут быть обнаружены при помощи микроскопа. К таким мутациям относятся утраты участков хромосом (делеции), добавление участков, поворот участка хромосомы на 180°, появление повторов.
- Геномные мутации вызвываются изменением числа хромосом. Могут появляться лишние гомологичные хромосомы: в хромосом ном наборе на месте двух гомологичных хромосом оказываются три -трисомия. В случае моносомии наблюдается утрата одной хромосомы из пары. При полиплоидии происходит кратное увеличение генома. Еще один вариант геномной мутации - гаплоидия, при которой остается только одна хромосома из каждой пары.
На частоту возникновения мутаций влияют, как уже было сказано, самые разнообразные факторы. При возникновении ряда геномных мутаций большое значение имеет, в частности, возраст матери.
Комбинативная изменчивость.
Данный тип изменчивости определяется характером полового процесса. При комбинативной изменчивости возникают новые генотипы из-за новых комбинаций генов. Этот тип изменчивости проявляется уже на стадии образования половых клеток. Как уже было сказано, в каждой половой клетке (гамете) представлена только одна гомологичная хромосома из каждой пары. Хромосомы попадают в гамету случайным образом, поэтому половые клетки одного человека могут довольно сильно отличаться по набору генов в хромосомах. Еще более важная стадия для возникновения комбинативной изменчивости - это оплодотворение, после которого у вновь возникшего организма 50% генов унаследовано от одного родителя, и 50% - от другого.
Модификационная изменчивость не связана с изменениями генотипа, а вызвана влиянием среды на развивающийся организм.
Наличие модификационной изменчивости очень важно для понимания сущности наследования. Наследуются не признаки. Можно взять организмы с абсолютно одинаковым генотипом, например вырастить черенки от одного и того же растения, но поместить их при этом в разные условия (освещенность, влажность, минеральное питание) и получить достаточно сильно отличающиеся растения с разными признаками (рост, урожайность, форма листьев и т. п.). Для описания реально сформировавшихся признаков организма используют понятие «фенотип».
Фенотип - это весь комплекс реально возникших признаков организма, который формируется как результат взаимодействия генотипа и влияний среды в ходе развития организма. Таким образом, сущность наследования заключается не в наследовании признака, а в способности генотипа в результате взаимодействия с условиями развития давать определенный фенотип.
Так как модификационная изменчивость не связана с изменениями генотипа, то модификации не передаются по наследству. Обычно это положение почему-то с трудом принимается. Кажется, что если, скажем, родители на протяжении нескольких поколений тренируются в поднятии тяжестей и обладают развитой мускулатурой, то эти свойства должны обязательно передаться детям. Между тем, это типичная модификация, а тренировки - это и есть то воздействие среды, которое повлияло на развитие признака. Никаких изменений генотипа при модификации не происходит и приобретенные в результате модификации признаки не наследуются. Дарвин называл этот вид изменчивости - ненаследственной.
Для характеристики пределов модификационной изменчивости применяется понятие норма реакции. Некоторые признаки у человека невозможно изменить за счет средовых влияний, например группу крови, пол, цвет глаз. Другие, напротив, очень чувствительны к воздействию среды. К примеру, в результате длительного пребывания на солнце цвет кожи становится темнее, а волосы светлеют. На вес человека сильно влияют особенности питания, болезни, наличие вредных привычек, стресс, образ жизни.
Средовые воздействия могут приводить не только к количественным, но и к качественным изменениям фенотипа. У некотррь« видов примулы при пониженной температуре воздуха(15-20 С) появляются цветы красного цвета, если же растения поместить во влажную среду с температурой 30°С, то образуются белые цветки.
причем, хотя норма реакции характеризует ненаследственную форму изменчивости (модификационную изменчивость), она тоже определяется генотипом. Это положение очень важно: норма реакции зависит от генотипа. Одно и то же воздействие среды на генотип может привести к сильному изменению одного его признака и никак не повлиять на другой.
21. Ген - функциональная единица наследственности. Молекулярное строение гена у прокариот и эукариот. Уникальные гены и повторы ДНК. Структурные гены. Гипотеза «1 ген- 1 фермент», её современная трактовка.
Ген - структурная и функциональная единица наследственности, контролирующая развитие определённого признака или свойства. Совокупность генов родители передают потомкам во время размножения. Термин «ген» был введён в употребление в 1909 году датским ботаником Вильгельмом Йогансеном. Изучением генов занимается наука генетика, родоначальником которой считается Грегор Мендель, который в 1865 году опубликовал результаты своих исследований о передачи по наследству признаков при скрещивании гороха. Гены могут подвергаться мутациям - случайным или целенаправленным изменениям последовательности нуклеотидов в цепи ДНК. Мутации могут приводить к изменению последовательности, а следовательно изменению биологических характеристик белка или РНК, которые, в свою очередь, могут иметь результатом общее или локальное изменённое или анормальное функционирование организма. Такие мутации в ряде случаев являются патогенными, так как их результатом является заболевание, или летальными на эмбриональном уровне. Однако, далеко не все изменения последовательности нуклеотидов приводят к изменению структуры белка (благодаря эффекту вырожденности генетического кода) или к существенному изменению последовательности и не являются патогенными. В частности, геном человека характеризуется однонуклеотидными полиморфизмами и вариациями числа копий, такими как делеции и дупликации, которые составляют около 1 % всей нуклеотидной последовательности человека. Однонуклеотидные полиморфизмы, в частности, определяют различные аллели одного гена.
У человека в результате делеции:
Синдром Вольфа- утрачен участок в большой хромосоме 4 ,
Синдром “кошачьего крика”- при делеции в хромосоме 5. Причина: хромосомная мутация потеря фрагмента хромосомы в 5-й паре.
Проявление: неправильное развитие гортани, крики, подобные кошачьим, I раннем детском возрасте, отставание в физическом и умственном развитии.
Мономеры, составляющие каждую из цепей ДНК, представляют собой сложные органические соединения, включающие в себя азотистые основания: аденин(А) или тимин(Т) или цитозин(Ц) или гуанин(Г), пятиатомный сахар-пентозу-дезоксирибозу, по имени которой и получила название сама ДНК, а также остаток фосфорной кислоты. Эти соединения носят название нуклеотидов.
Хромосома любого организма, будь то бактерия или человек, содержит длинную непрерывную цепь ДНК. вдоль которой расположено множество генов. Различные организмы резко отличаются по количеству ДНК, составляющей их геномы. У вирусов в зависимости от их величины и сложности размер генома колеблется от нескольких тысяч до сотен пар нуклеотидов. Гены в таких просто устроенных геномах расположены один за другим и занимают до 100% длины соответствующей нуклеиновой кислоты (РНК и ДНК). Для многих вирусов установлена полная нуклеотидная последовательность ДНК. У бактерий размер генома значительно больше. У кишечной палочки единственная нить ДНК – бактериальная хромосома состоит из 4,2х106(6 степень) пар нуклеотидов. Более половины этого количества состоит из структурных генов, т.е. генов, кодирующих определенные белки. Остальную часть бактериальной хромосомы составляют неспособные транскрибироваться нуклеотидные последовательности, функция которых не вполне ясна. Подавляющее большинство бактериальных генов уникальны, т.е. представлены в геноме один раз. Исключение составляют гены транспортных и рибосомальных РНК, которые могут повторяться десятки раз.
Геном эукариот, особенно высших, резко превышает по размерам геном прокариот и достигает, как отмечалось, сотен миллионов и миллиардов пар нуклеотидов. Количество структурных генов при этом возрастает не очень сильно. Количество ДНК в геноме человека достаточно для образования примерно 2 млн. структурных генов. Реально имеющееся число оценивается как 50-100 тыс. генов, т.е. в 20-40 раз меньше того, что могло бы кодироваться геномом такого размера. Следовательно, приходится констатировать избыточность генома эукариот. Причины избыточности в настоящее время в значительной степени прояснились: во-первых, некоторые гены и последовательности нуклеотидов многократно повторены, во-вторых, в геноме существует много генетических элементов, имеющих регуляторную функцию, в-третьих, часть ДНК вообще не содержит генов.
Согласно современным представлениям, ген, кодирующий синтез определенного белка, у эукариот состоит из нескольких обязательных элементов. Прежде всего это обширная регуляторная зона, оказывающая сильное влияние на активность гена в той или иной ткани организма на определенной стадии его индивидуального развития. Далее расположен непосредственно примыкающий к кодирующим элементам гена промотор – последовательность ДНК длиной до 80-100 пар нуклеотидов, ответственная за связывание РНК-полимеразы, осуществляющей транскрипцию данного гена. Вслед за промотором лежит структурная часть гена, заключающая в себе информацию о первичной структуре соответствующего белка. Эта область для большинства генов эукариот существенно короче регуляторной зоны, однако ее длина может измеряться тысячами пар нуклеотидов.
Важная особенность эукариотических генов – их прерывность. Это значит, что область гена, кодирующая белок, состоит из нуклеотидных последовательностей двух типов. Одни – экзоны – это участки ДНК, которые несут информацию и строении белка и входят в состав соответствующих РНК и белка. Другие – интроны – не кодируют структуру белка и в состав зрелой молекулы и-РНК не входят, хотя и транскрибируются. Процесс вырезания интронов – «ненужных» участков молекулы РНК и сращивания экзонов при образовании и-РНК осуществляется специальными ферментами и получил название Сплайсинг (сшивание, сращивание).
Геном эукариот характеризуется двумя основными особенностями:
1) Повторенность последовательностей
2) Разделением по составу на различные фрагменты, характеризуемые специфическим содержанием нуклеотидов
Повторенная ДНК состоит из нуклеотидных последовательностей различной длины и состава, которые встречаются в геноме несколько раз либо в тандемно-повторенном, либо в диспергированном виде. Последовательности ДНК, которые не повторяются, называются уникальной ДНК. Размер части генома, занятой повторяющимися последовательностями, широко варьирует между таксонами. У дрожжей он достигает 20%, у млекопитающих до 60% всей ДНК повторяется. У растений процент повторенных последовательностей может превышать 80%.
По взаимной ориентации в структуре ДНК различаются прямые, инвертированные, симметричные повторы, палиндромы, комплементарные палиндромы и т.п. В очень широком диапазоне варьирует и длина (в числе оснований) элементарной повторяющейся единицы, и степень их повторяемости, и характер распределения в геноме. периодичность повторений ДНК может иметь очень сложную структуру, когда короткие повторы включены в более протяженные или окаймляют их и т.д. Кроме того, для последовательностей ДНК можно рассматривать зеркальные и инвертированные повторы. Геном человека известен на 94%.На основании этого материала можно сделать следующий вывод- повторы занимают по крайней мере 50% генома.
СТРУКТУРНЫЕ ГЕНЫ - гены, кодирующие клеточные белки с ферментативными или структурными функциями. К ним же относят гены, кодирующие структуру рРНК и тРНК. Есть гены, содержащие информацию о структуре полипептидной цепи, в конечном счете – структурных белках. Такие последовательности нуклеотидов длинной в один ген, называются структурными генами. Гены, определяющие место, время, длительность включения структурных генов – регуляторные гены.
Гены имеют маленький размер, хотя состоят из тысяч пар нуклеотидов. Наличие гена устанавливается по проявлению признака гена (конечному продукту). Общую схему строения генетического аппарата и его работы в 1961 году предложили Жакоб, Моно. Они предложил, что есть участок молекулы ДНК с группой структурных генов. К этой группе примыкает участок в 200пар нуклеотидов – промотор (участок примыкания ДНК зависимой РНК-полимеразы). К этому участку примыкает ген-оператор. Название всей системы – оперон. Регуляция осуществляется регуляторным геном. В итоге белок-репрессор взаимодействует с геном-оператором, и оперон начинает работать. Субстрат взаимодействует с геном регуляторами, оперон блокируется. Принцип обратной связи. Экспрессия оперона включается как единое целое. 1940 год - Бидл и Татум предложили гипотезу: 1 ген – 1 фермент. Эта гипотеза сыграла важную роль – ученые стали рассматривать конечные продукты. Оказалось, что гипотеза имеет ограничения, т.к. все ферменты – белки, но не все белки – ферменты. Как правило, белки являются олигомерами – т.е. существуют в четвертичной структуре. Пример, капсула табачной мозаики имеет более 1200 полипептидов. У эукариот экспрессия (проявление) генов не исследована. Причина – серьезные препятствия:
Организация генетического материала в форме хромосом
У многоклеточных организмов клетки специализированы и поэтому часть генов выключена.
Наличие гистоновых белков, в то время как у прокариот - «голая» ДНК.
Гистоновые и негистоновые белки принимают участие в экспрессии генов, участвуют в создании структуры.
22. Классификация генов: гены структурные, регуляторы. Свойства генов (дискретность, стабильность, лабильность, полиаллелизм, специфичность, плейотропия).
Дискретность - несмешиваемость генов
Стабильность - способность сохранять структуру
Лабильность - способность многократно мутировать
Множественный аллелизм - многие гены существуют в популяции во множестве молекулярных форм
Аллельность - в генотипе диплоидных организмов только две формы гена
Специфичность - каждый ген кодирует свой признак
Плейотропия - множественный эффект гена
Экспрессивность - степень выраженности гена в признаке
Пенетрантность - частота проявления гена в фенотипе
Амплификация - увеличение количества копий гена.

23. Строение гена. Регуляция экспрессия генов у прокариот. Гипотеза оперона.
Экспрессия генов - это процесс, в ходе которого наследственная информация от гена (последовательности нуклеотидов ДНК) преобразуется в функциональный продукт - РНК или белок. Экспрессия генов может регулироваться на всех стадиях процесса: и во время транскрипции, и во время трансляции, и на стадии посттрансляционных модификаций белков.
Регуляция экспрессии генов позволяет клеткам контролировать собственную структуру и функцию и является основой дифференцировки клеток, морфогенеза и адаптации. Экспрессия генов является субстратом для эволюционных изменений, так как контроль за временем, местом и количественными характеристиками экспрессии одного гена может иметь влияние на функции других генов в целом организме. У прокариот и эукариот гены представляют собой последовательности нуклеотидов ДНК. На матрице ДНК происходит транскрипция - синтез комплементарной РНК. Далее на матрице мРНК происходит трансляция - синтезируются белки. Существуют гены, кодирующие нематричную РНК (например, рРНК, тРНК, малые РНК), которые экспрессируются (транскрибируются), но не транслируются в белки.
Исследования на клетках Е. coli позволили установить, что у бактерий существуют ферменты 3 типов:
конститутивные, присутствующие в клетках в постоянных количествах независимо от метаболического состояния организма (например, ферменты гликолиза)
индуцируемые, их концентрация в обычных условиях мала, но может возрастать в 100Q раз и более, если, например, в среду культивирования клеток добавить субстрат такого фермента
репрессируемые, т.е. ферменты метаболических путей, синтез которых прекращается при добавлении в среду выращивания конечного продукта этих путей.
На основании генетических исследований индукции β-галактозидазы, участвующей в клетках Е. coli, в гидролитическом расщеплении лактозы Франсуа Жакоб и Жак Моно в 1961 г. сформулировали гипотезу оперона, которая объясняла механизм контроля синтеза белков у прокариотов.
В экспериментах гипотеза оперона получила полное подтверждение, а предложенный в ней тип регуляции стали называть контролем синтеза белка на уровне транскрипции, так как в этом случае изменение скорости синтеза белков осуществляется за счёт изменения скорости транскрипции генов, т.е. на стадии образования мРНК.
У Е. coli, как и у других прокариотов, ДНК не отделена от цитоплазмы ядерной оболочкой. В процессе транскрипции образуются первичные транскрипты, не содержащие нитронов, а мРНК лишены "кэпа" и поли-А-конца. Синтез белка начинается до того, как заканчивается синтез его матрицы, т.е. транскрипция и трансляция протекают почти одновременно. Исходя из размера генома (4×106 пар нуклеотидов), каждая клетка Е. coli содержит информацию о нескольких тысячах белков. Но при нормальных условиях роста она синтезирует около 600-800 различных белков, а это означает, что многие гены не транскрибируются, т.е. неактивны. Гены белков, функции которых в метаболических процессах тесно связаны, часто в геноме группируются вместе в структурные единицы (опероны). Согласно теории Жакоба и Моно, оперонами называют участки молекулы ДНК, которые содержат информацию о группе функционально взаимосвязанных структурных белков, и регуляторную зону, контролирующую транскрипцию этих генов. Структурные гены оперона экспрессируются согласованно, либо все они транскрибируются, и тогда оперон активен, либо ни один из генов не "прочитывается", и тогда оперон неактивен. Когда оперон активен и все его гены транскрибируются, то синтезируется полицистронная мРНК, служащая матрицей для синтеза всех белков этого оперона. Транскрипция структурных генов зависит от способности РНК-полимеразы присоединяться к промотору, расположенному на 5"-конце оперона перед структурными генами.
Связывание РНК-полимеразы с промотором зависит от присутствия белка-репрессора на смежном с промотором участке, который называют "оператор". Белок-репрессор синтезируется в клетке с постоянной скоростью и имеет сродство к операторному участку. Структурно участки промотора и оператора частично перекрываются, поэтому присоединение белка-репрессора к оператору создаёт стерическое препятствие для присоединения РНК-полимеразы.
Большинство механизмов регуляции синтеза белков направлено на изменение скорости связывания РНК-полимеразы с промотором, влияя таким образом на этап инициации транскрипции. Гены, осуществляющие синтез регуляторных белков, могут быть удалены от оперона, транскрипцию которого они контролируют.
Биохакер Джошуа Зайнер хочет создать мир, в котором любой человек способен и в праве экспериментировать со своей ДНК. Почему нет?
«У нас здесь немного ДНК и шприц», – говорит Джошуя Зайнер в комнате, полной синтетических биологов и других исследователей. Он наполняет иглу и вонзает её в кожу. «Это изменит мои мышечные гены, и даст мне больше мышечной массы».
Зайнер – биохакер, он экспериментирует с биологией в DIY, а не в обычной лаборатории, – выступил на конференции SynBioBeta в Сан-Франциско с докладом «Пошаговое руководство по генетическому изменению себя с помощью CRISPR», где в других презентациях участвовали академики в костюмах и молодые руководители типичных биотехнологических стартапов. В отличие от прочих, он начал своё выступление, раздавая образцы и буклеты, в которых объяснялись основы DIY генной инженерии.
Биохакер Зайнер выступил на конференции SynBioBeta с докладом «Пошаговое руководство по генетическому изменению себя с помощью CRISPR»
Если вы хотите генетически модифицировать себя, – это не обязательно сложно. Когда он предложил образцы в маленьких пакетиках толпе, Зайнер объяснил, что ему потребовалось около пяти минут, чтобы сделать ДНК, которую он привёз на презентацию. В пробирке был Cas9, фермент, который разрезает ДНК в определённом месте, ориентированном по направляющей РНК, в системе редактирования генов, известной как CRISPR. В этом примере он был разработан для выключения гена миостатина, который вырабатывает гормон, ограничивающий рост мышц и уменьшающий мышечную массу. В исследовании, проведённом в Китае, собаки с отредактированным геном имели удвоенную мышечную массу. Если кто-то из зрителей захотел попробовать, они могли взять пробирку домой и ввести его позже. Даже капая его на кожу, сказал Зейнер, вы получите эффект, хотя и ограниченный.
Зайнер имеет докторскую степень по молекулярной биологии и биофизике, он также работал научным сотрудником в НАСА по модификации организмов для жизни на Марсе. Но он полагает, что синтетическая биология для редактирования других организмов или себя может стать столь же простой в использовании, как, например, CMS для создания веб-сайта.
«Вам не нужно знать, какой промотор использовать чтобы заставить работать нужный ген или фрагмент ДНК, – говорит он, используя некоторые технические термины из генной инженерии. «Вы не хотите знать, какой терминатор использовать, или ориджин репликации… Инженер, программирующий ДНК, должен знать, как это сделать. Но единственное, что вам нужно знать, – так, я хочу, чтобы гриб был фиолетовым. Это не должно быть сложнее. Всё это вполне возможно – это просто создание инфраструктуры и платформы, чтобы любой мог это сделать».
Конечно, магазин приложений для генетического редактирования ещё не создан. Но немалое число биохакеров узнали достаточно, чтобы – порой необдуманно – экспериментировать над собой. Несколько человек, которых Зайнер знает, например, начали вводить себе миостатин. «Это происходит прямо сейчас», – говорит он. «Все эти вещи начали появляться буквально в последние несколько недель». Пока ещё рано говорить о том, улучшили ли инъекции экспериментаторов или вызвали проблемы, но некоторые надеются увидеть результаты в ближайшие месяцы.
Несмотря на проведённое в академических кругах время, Зайнер явно не является типичным исследователем и избегает идеи, что эксперименты должны ограничиваться лабораториями. Когда в НАСА он начал общаться с другими биохакерами через список рассылки, и узнал о проблемах тех, кто хотел делать DIY работу, – поставщиков было трудно найти, и они не всегда отправляли нужные заказы тем, у кого не было лаборатории, – он в 2013 году начал бизнес под названием The ODIN (Open Discovery Institute, and an homage to the Norse god), чтобы пересылать комплекты и инструменты людям, желающим работать в своём гараже или комнате. В 2015 году, решив покинуть НАСА, потому что ему не нравилось работать в их консервативной среде, он запустил успешную кампанию по сбору средств для набора DIY CRISPR.
«Единственное, что вам нужно знать, – так, я хочу, чтобы гриб был фиолетовым. Это не должно быть сложнее.»
В 2016 году он продал продуктов на сумму $200 000, включая набор для дрожжей, которые можно использовать для заправки светящегося биолюминесцентного пива, набор для обнаружения антибиотиков дома и полную домашнюю лабораторию по цене MacBook Pro. В 2017 году он ожидает удвоения продаж. Многие наборы просты, и большинство покупателей, вероятно, не используют их, чтобы изменить себя (многие комплекты идут в школы). Но Зайнер также надеется, что по мере получения новых знаний, люди будут экспериментировать более необычными способами.
Зайнер продает полную домашнюю лабораторию биохакинга примерно по цене MacBook Pro.
Он задаёт вопрос, являются ли традиционные методы исследования, такие как рандомизированные контролируемые испытания, единственным способом сделать открытия, указывая на то, что в новой персонализированной медицине (такой как иммунотерапия рака, которая персонализирована для каждого пациента), размер выборки одного человека имеет смысл. В своём выступлении он утверждал, что люди должны иметь возможность самостоятельно экспериментировать, если захотят; мы меняем нашу ДНК, когда пьём алкоголь или курим сигареты, или дышим грязным городским воздухом. Многие действия, санкционированные обществом, более опасны. «Мы жертвуем, возможно, миллион людей в год автомобильным богам» – сказал он. «Если вы спросите кого-нибудь: "Не могли бы вы избавиться от автомобилей?" – нет.» (Зайнер экспериментировал по-разному, включая экстремальную пересадку фекалий DIY, которая, по его словам, вылечила его проблемы с пищеварением, он также помогает больным раком с DIY иммунотерапией).
Если вы измените свою ДНК, вы можете затем секвенировать свой геном, чтобы увидеть, произошло ли изменение. Но эксперимент в гараже не может предоставить столько информации, сколько обычные методы. «Вы можете подтвердить, что вы изменили ДНК, но это не значит, что оно безопасно и эффективно», – говорит Джордж Чёрч, профессор генетики в Гарвардской Медицинской Школе (которая также выступает в качестве адвайзера компании Зайнера, признавая ценность биологически грамотной публики в веке биологии). «Всё, что он делает, – говорит вам, что вы сделали правильную работу, но это может быть опасно, потому что вы также изменили что-то ещё. Это может быть неэффективным в том смысле, что недостаточно клеток было изменено, или уже слишком поздно, и ущерб уже был нанесён». Если ребёнок рождается с микроцефалией, например, изменение генов в его теле, скорее всего, не повлияет на его мозг.
«Мы живём в невероятное время, когда мы изучаем очень многое в биологии и генетики благодаря CRISPR, но мы всё ещё многого не знаем о безопасности редактирования человеческих клеток с помощью CRISPR.»
Любой, кто хочет ввести себе модифицированную ДНК, рискует без достаточного количества данных или, возможно, любых реальных данных – о том, что может произойти, для принятия обоснованного решения. Это, наверное, само собой разумеется: не пытайтесь делать это дома. «Мы живём в невероятное время, когда мы изучаем очень многое в биологии и генетики благодаря CRISPR, но мы всё ещё многого не знаем о безопасности редактирования человеческих клеток с помощью CRISPR», – говорит Алекс Марсон, исследователь в области микробиологии и иммунологии в Калифорнийском университете в Сан-Франциско и эксперт по CRISPR. «Очень важно, чтобы оно проходило через тщательные и проверенные тесты безопасности в каждом случае, и делалось ответственным образом».
В Германии биохакинг теперь вне закона, и человек, проводящий эксперименты за пределами лицензированной лаборатории, может получить штраф в размере €50 000 или три года в тюрьме. Всемирное антидопинговое агентство теперь запрещает все формы редактирования генов у спортсменов. Однако в США биохакинг ещё не регулируется. И Зайнер не считает, что вообще должен, он сравнивает опасения, что люди изучают, как использовать синтетическую биологию, с опасениями изучения, как использовать компьютеры, в начале 1980-х. (Он приводит интервью 1981 года, в котором Тед Коппел спросил Стива Джобса, есть ли опасность того, что люди окажутся под контролем компьютеров.) Зайнер надеется продолжать помогать как можно большему числу людей, становится более «грамотными в ДНК».
«Я хочу жить в мире, где люди генетически модифицируют себя. Я хочу жить в мире, где все эти классные вещи, которые мы видим в научно-фантастических телешоу, реальны. Может быть, я сумасшедший и глупый… но я думаю, наверное, это действительно возможно».
Вот почему, он сделал себе инъекцию перед толпой на конференции. «Я хочу, чтобы люди перестали спорить о том, можно ли использовать CRISPR или нельзя, нормально ли генетически модифицировать себя», – говорит он. «Уже слишком поздно: я сделал выбор за вас. Споры закончены. Давайте продолжим. Давайте использовать генную инженерию, чтобы помочь людям. Или дать им фиолетовую кожу».
Мутация (лат. mutatio - изменение) - стойкое преобразование генотипа, происходящее под влиянием внешней или внутренней среды.
Геномные мутации - это мутации, которые приводят к добавлению либо утрате одной, нескольких или полного гаплоидного набора хромосом. Разные виды геномных мутаций называют гетероплоидией и полиплоидией.
Полиплоидия – кратные изменения (в несколько раз, например, 12 → 24). У животных не встречается, у растений приводит к увеличению размера.
Анеуплоидия – изменения на одну-две хромосомы. Например, одна лишняя двадцать первая хромосома приводит к синдрому Дауна (при этом общее количество хромосом – 47)
26.Изменение числа и порядка расположения генов (хромосомные перестройки)
Хромосомные перестройки (их также называют аберрациями) возникают в случае двух или более хромосомных разрывов.
· Делеция , или нехватка. Утрачен участок хромосомы.
· Дупликация , или удвоение. Один из участков хромосомы представлен в хромосомном наборе более одного раза.
· Инверсия возникает в результате двух разрывов в одной хромосоме, но при условии, что внутренний фрагмент хромосомы совершит поворот на 180 градусов, т.е. его полярность измениться на обратную.
Инвертированный участок хромосомы может включать или не включать центромеру. В первом случае инверсия называется перицентрической (т.е. охватывающей центромеру), а во втором –парацентрической (околоцентромерной).
Транслокации . Если разрывы оказываются в двух хромосомах, то при воссоединении возможен обмен фрагментами. При симметричном воссоединении образуются новые хромосомы, в которых произошел обмен дистальными участками негомологичных хромосом. Такие транслокации называются реципрокными.
Участок хромосомы может также изменять свое положение и без реципрокного обмена, оставаясь в той же хромосоме, или включаясь в какую-нибудь другую. Такие нереципрокные транслокации иногда называют транспозициями .
В случае соединения двух акроцентрических хромосом в районе их центромер с потерей коротких плеч наблюдается центрическое слияние –робертсоновская транслокация.
27.Изменение индивидуальных генов (генная мутация)
Мутации (от лат. mutatio - перемена) - это изменение генов и хромосом, фенотипически проявляющиеся в изменении свойств и признаков организмов.
Генные (точковые) мутации - это изменения числа и/или последовательности нуклеотидов в структуре ДНК (вставки, выпадения, перемещения, замещения нуклеотидов) в пределах отдельных генов, приводящие к изменению количества или качества соответствующих белковых продуктов.
Передача генной мутации.
Она происходит по обычным законам наследственности. Риск для потомства более или менее велик в зависимости от того, доминантным или рецессивным является «больной» ген и где он находится - в обычной хромосоме или в половой. Имейте только в виду, что в случае, если ген оказывается рецессивным, человек может передать его своему потомству.
Типичным примером является гемофилия - болезнь крови (нарушение ее свертываемости). Эта болезнь отличается тем, что она передается только женщинами, но вызывает нарушения только у мужчин; другими словами, женщина, внешне здоровая, может перечить эту болезнь одному из своих сыновей.
Изменения структурных генов.
1) «Сдвиг рамки считывания» - вставка или выпадение пары или нескольких пар нуклеотидов. Например, исходный порядок нуклеотидов: АГГАЦТЦГА..., а после вставки нуклеотида: ААГГАЦТЦГА...; в зависимости от места вставки или выпадения нуклеотидов изменяется меньшее или большее число кодонов.
2) Транзиция - замена оснований: пуринового на пуриновое, или пиримидинового на пиримидиновое; например: А ↔ Г, Ц ↔ Т; при этом изменяется тот кодон, в котором произошла транзиция.
3) Трансверзия - замена пуринового основания на пиримидиновое или пиримидинового на пуриновое. Например: А ↔ Ц, Г ↔ Т; изменяется тот кодон, в котором произошла трансверзия.
Изменения структурных генов приводят к: а) мисценс-мутациям - замене нуклеотида в кодирующей части гена, приводящей к замене аминокислоты в полипептиде; б) нонсенс-мутациям - образованию «бессмысленных» кодонов (УАА, УАГ, УГА), не кодирующих аминокислоты (терминаторы, определяющие окончание считывания), и прекращению трансляции.
1) Белок-репрессор «не подходит» к гену-оператору («ключ не входит в замочную скважину») - структурные гены работают постоянно (белки синтезируются все время).
2) Белок-репрессор плотно «присоединяется» к гену-оператору и не снимается индуктором («ключ не выходит из замочной скважины») - структурные гены постоянно не работают, и не синтезируются белки, закодированные в данном транскриптоне.
3) Нарушение чередования репрессии и индукции - при отсутствии индуктора специфический белок синтезируется, а при его наличии – не синтезируется. Вышеназванные нарушения работы транскриптонов связаны с мутациями гена-регулятора или гена-оператора.
Некоторые мутации не оказывают влияния на функцию соответствующего полипептида. Значительная часть генных мутаций нарушает функционирование гена и приводит к развитию генных болезней. Фенотипически генные мутации проявляются признаками нарушений метаболизма, частота которых в популяциях человека составляет 1-2% (фенилкетонурия, гемофилии, нейрофиброматоз, муковисцидоз, мышечная дистрофия Дюшенна-Беккера, гемоглобинопатия S). Они выявляются биохимическими методами и методами рекомбинантной ДНК.
По последствиям генные мутации классифицируют на нейтральные , регуляторные и динамические .
Нейтральная (молчащая) мутация - не имеет фенотипического выражения (например, замена нуклеотидов, не приводящая к замене аминокислоты в силу вырожденности генетического кода).
Регуляторная мутация - мутация в 5"- или З" - нетранслируемых областях гена; нарушает экспрессию гена.
Динамические мутации - обусловлены увеличением числа тринуклеотидных повторов в функционально значимых частях гена. Такие мутации могут привести к торможению или блокаде транскрипции, приобретению белковыми молекулами свойств, нарушающих их нормальный метаболизм.
ХРОМОСОМНЫЕ МУТАЦИИ
На хромосомном уровне организации наследственный материал обладает всеми характеристиками субстрата наследственности и изменчивости, в том числе способностью к приобретению изменений, которые могут передаваться новому поколению . Под влиянием различных воздействий физико-химическая и морфологическая структура хромосом может изменяться. Разрывы хромосом происходят закономерно в ходе кроссинговера , когда гомологичные хромосомы обмениваются соответствующими участками. Нарушение кроссинговера, при котором хромосомы обмениваются неравноценным генетическим материалом, приводит к появлению новых групп сцепления с изменением числа генов. При хромосомных мутациях последовательность нуклеотидов в генах обычно не меняется, но изменение числа или положения генов может привести к генетическому дисбалансу, что пагубно сказывается на нормальном развитии организма.
Хромосомные аберрации могут быть внутрихромосомными , межхромосомными и изохромосомными .
К внутрихромосомным относятся перестройки внутри одной хромосомы.
1) Делеции (нехватки) - выпадение части хромосомы (утрата сегмента ДНК размером от одного нуклеотида до целых генов) – может стать причиной нарушения эмбриогенеза и формирования множественных аномалий развития. Например, делеция участка короткого плеча 5-ой (5р-) хромосомы у человека приводит к развитию синдрома кошачьего крика (недоразвитие гортани, ВПР сердца, отставание умственного развития; у больных детей из-за аномалии гортани плач напоминает кошачье мяуканье). При делеции теломеров обоих плеч хромосомы часто наблюдается замыкание оставшейся структуры в кольцо - кольцевые хромосомы . При выпадении центромерного участка образуются децентрические хромосомы .
2) Дупликация - удвоение участка хромосомы. Результатом дупликации во второй хромосоме мухи дрозофилы может служить появление полосковидных глаз. Например, трисомия по короткому плечу хромосомы 9 приводит к появлению множественных ВПР, включая микроцефалию, задержку физического, психического и интеллектуального развития.
3) Инверсия - отрыв фрагмента хромосомы, поворот его на 180° и прикрепление к месту отрыва. При этом наблюдается нарушение порядка расположения генов.
4) Инсерция - вставка фрагментов ДНК размером от одного нуклеотида до целого гена.
5) Транспозиция - присоединение фрагмента к своей же хромосоме, но в другом месте.
Делеции и дупликации проявляются фенотипически всегда, так как изменяется набор генов и наблюдаются частичные моносомии (моносомии по части хромосомы) при нехватках и частичные трисомии - при дупликациях. Инверсии и транслокации фенотипически проявляются не всегда; они могут быть сбалансированными, когда не происходит ни увеличения, ни уменьшения генетического материала и сохраняется общий баланс генов в геноме. При инверсиях и транслокациях затрудняется конъюгация гомологичных хромосом, что может служить причиной нарушения при распределении генетического материала между дочерними клетками.
Межхромосомные перестройки происходят между негомологичными хромосомами.
Транслокация - это обмен сегментами между негомологичными хромосомами. Различают транслокации:
- реципрокные , когда две хромосомы обмениваются сегментами;
- нереципрокные , когда сегменты одной хромосомы переносятся в другую;
- робертсоновские , когда две акроцентрические хромосомы соединяются своими центромерными районами с потерей коротких плеч; в результате образуется одна метацентрическая хромосома.
Изохромосомные аберрации - образование одинаковых, но зеркальных фрагментов двух разных хромосом, содержащих одни и те же наборы генов. Это происходит в результате поперечного разрыва хроматид через центромеры (отсюда другое название - центрическое соединение).
Хромосомные аберрации выявляются цитогенетическими методами.
ГЕНОМНЫЕ МУТАЦИИ
Геномные мутации характеризуются изменением числа хромосом. Причиной геномных мутацийможет быть нарушение процессов, протекающих в мейозе. Нарушение расхождения бивалентов в анафазе приводит к появлению гамет с разным количеством хромосом (на каждую гамету с лишней хромосомой приходится другая - без одной хромосомы). Оплодотворение таких гамет нормальными половыми клетками приводит к изменению общего числа хромосом в кариотипе за счет уменьшения (моносомия ) или увеличения (трисомия ) числа отдельных хромосом.
Анеуплоидия - некратное гаплоидному уменьшение или увеличение числа хромосом (2n±1, 2n±2 и т. д.). Разновидности анеуплоидии: а) трисомия - три гомологичных хромосомы в кариотипе, например, при синдроме Дауна (трисомия по 21-й хромосоме), синдроме Эдвардса (трисомия по 18-й паре хромосом), синдроме Патау (трисомия по 13-й паре); б) моносомия - в наборе только одна из пары гомологичных хромосом. Единственная совместимая с жизнью моносомия у человека – по хромосоме Х – приводит к развитию синдрома Шерешевского-Тернера (45,Х0). Моносомии по первым крупным парам хромосом являются для человека летальными мутациями. Нулисомия - отсутствие пары хромосом (летальная мутация).
При повреждении механизма распределения гомологичных хромосом клетка остается неразделившейся, и образуются диплоидные гаметы. Оплодотворение таких гамет приводит к увеличению числа наборов хромосом.
Полиплоидия - увеличение числа наборов хромосом, кратное гаплоидному (Зn, 4n, 5n, ...). Причины: двойное оплодотворение и отсутствие первого мейотического деления. Полиплоидия, как правило, используется в селекции растений и приводит к повышению урожайности. У млекопитающих и человека полиплоидия, а также большинство анеуплоидий, приводят к формированию леталей.
Гаплоидия (1n) - одинарный набор хромосом - например, у трутней пчел. Жизнеспособность гаплоидов снижается, так как в данном случае проявляются все рецессивные гены, содержащиеся в единственном числе. Для млекопитающих и человека - летальная мутация.
Геномные мутации обнаруживаются цитогенетическими методами. Они всегда проявляются фенотипически.
Патологические эффекты хромосомных и геномных мутаций проявляются на всех стадиях онтогенеза, поскольку вызывают нарушения:
Общего генетического баланса;
Скоординированности в работе генов;
Системности регуляции.
Они проявляются в двух взаимосвязанных вариантах:
Летальность;
Врожденные пороки развития.
Летальный исход хромосомных мутаций - один из главных факторов внутриутробной гибели, достаточно частой у человека. Многочисленные цитогенетические исследования материала спонтанных абортов, выкидышей и мертворожденных позволяют объективно судить об эффектах разных типов хромосомных аномалий во внутриутробном периоде индивидуального развития. Суммарный вклад хромосомных мутаций в число случаев внутриутробной гибели у человека составляет 45%. Среди перинатально погибших плодов частота хромосомных аномалий составляет 6%. В этих случаях летальные эффекты сочетаются с пороками развития, а точнее, реализуются через пороки.
Практически все хромосомные аномалии ведут к врожденным порокам развития . Более тяжелые их формы приводят к более раннему прерыванию беременности. Роль хромосомных и геномных мутаций не ограничивается только их влиянием на развитие патологических процессов в ранних периодах онтогенеза. Их эффекты прослеживаются в течение всей жизни.
В постнатальном периоде хромосомные мутации возникают в соматических клетках постоянно с невысокой частотой (около 2%). Такие клетки содержат специфические для них белки и в нормеэлиминируются иммунной системой, если они проявляют себя чужеродно. Однако в некоторых случаях хромосомные аномалии являются причиной злокачественного роста.
Облучение и химические мутагены, индуцирующие хромосомные аберрации, вызывают гибель клеток и, тем самым, способствуют развитию лучевой болезни, аплазии костного мозга. Имеются экспериментальные доказательства накопления клеток с хромосомными аберрациями в процессе старения.
ГЕНЕТИЧЕСКИЕ КОНЦЕПЦИИ КАНЦЕРОГЕНЕЗА
В настоящее время установлено, что при канцерогенезе изменения происходят на молекулярно-генетическом уровне и затрагивают механизмы, отвечающие за размножение, рост и дифференцировку клеток. Хронологически можно выделить несколько теорий (концепций) канцерогенеза.
1. Мутационная концепция. Впервые Г. де Фриз (1901) высказал предположение о том, что опухоли - это результат мутаций соматических клеток. Т. Бовери (1914) считал, что в основе канцерогенеза лежат геномные или хромосомные мутации. В дальнейшем было показано, что процесс канцерогенеза может происходить без структурных изменений в геноме, а обнаруживаемые в опухолевых клетках хромосомные и геномные мутации являются следствием перерождения клеток.
2. Вирусо-генетическая концепция. Еще в 1911 г. Ф. Раус впервые показал, что вирусы являются причиной саркомы кур. Затем была установлена вирусная природа некоторых других опухолей (лейкозов у кур, мышей и крыс, бородавок у человека и кроликов). Л.А. Зильбер (1944-1968) считал вирусы универсальной причиной злокачественного роста. Мутагены и канцерогены стимулируют активность вирусов, их геном включается в ДНК клетки и изменяет ее свойства.
3. Эпигеномная концепция. Ю.М. Оленов (1967) и А.Ю. Броновицкий (1972) считали, что в основе превращения нормальной клетки в опухолевую лежат стойкие нарушения регуляции генной активности, т.е. повреждаются функциональные гены.
4. Концепция онкогена. Р. Хюбнер (1969) и Г.И. Абелев (1975) объединили вторую и третью концепции. ДНК клеток любого организма содержит определенные участки - протоонкогены. Протоонкогены организм получает от предков, или их вносят интегративные вирусы. Они длительное время могут находиться в неактивном (репрессированном) состоянии. Активирование протоонкогенов может вызвать их мутация, внесение в клетку промотора вируса и др. Они преобразуются в онкогены, которые детерминируют синтез трансформирующих белков, превращающих нормальную клетку в опухолевую.
УРОВНИ ПАТОГЕНЕЗА НАСЛЕДСТВЕННЫХ БОЛЕЗНЕЙ
Основное звено патогенеза для многих наследственных заболеваний находится на клеточном уровне , где происходят патологические процессы, характерные для данной конкретной нозологической формы. Точкой приложения первичного действия мутантного гена являются отдельные структуры клетки, разные при различных болезнях: лизосомы, пероксисомы, мембраны, митохондрии.
На лизосомном уровне развертываются патогенетические процессыпри болезнях накопления в связи с нарушением активности лизосомных ферментов. Так, накопление в клетках, а затем и в основном межклеточном веществе, гликозаминогликанов (мукополисахаридов) приводит к развитию тяжелых заболеваний - мукополисахаридозов. Причина избыточного содержания этих полимеров - отсутствие их деградации в лизосомах, что связано с дефектами в группе специфических ферментов, катализирующих весь цикл деградации.
Если точкой приложения действия мутантного гена становятся пероксисомы, то развиваются пероксисомные болезни . Клинически эти заболевания проявляются в виде множественных врожденных пороков развития , в целом сходных при разных нозологических формах (множественные черепно-лицевые дисморфии, катаракта, почечные кисты и другие проявления).
Клеточные мембраны тоже могут быть ключевыми элементами патогенеза генных болезней. Так, отсутствие специфических белковых молекул-рецепторов на клеточной поверхности, связывающих липопротеиды низкой плотности, приводит к семейной гиперхолестеринемии
Клеточный уровень патогенеза генных болезней может проявляться не только в конкретных органеллах, но и в виде нарушений скоординированности деятельности клетки. Например, мутации, затрагивающие области онкогенов, ведут к снятию контроля размножения клеток и, соответственно, к злокачественному росту.
На органном уровне при различных болезнях мишенью патологического процесса служат различные органы. Поражение их может быть первичным или вторичным:
Отложение меди в печени и экстрапирамидной системе мозга при гепатолентикулярной дегенерации (болезнь Вильсона-Коновалова) - первичный процесс;
Гемосидероз паренхиматозных органов при первичном гемохроматозе или талассемии развивается вторично вследствие усиленного распада эритроцитов.
КЛАССИФИКАЦИЯ НАСЛЕДСТВЕННЫХ БОЛЕЗНЕЙ
1. По виду клеток, в которых произошли мутации: а) гаметические; б) соматические.
2. По клиническим проявлениям: а) предрасположение к болезни; б) собственно болезни; в) особые формы болезни – уродства.
3. По виду наследственности и аномального гена: а) доминантные; б) рецессивные; в) полудоминантные.
4. Наиболее практичной является классификация в зависимости от уровня организации наследственного материала: а) генные; б) хромосомные.
В зависимости от вида первично поражённых клеток выделены следующие группы заболеваний:
болезни вследствие мутаций в половых клетках - «гаметические», т.е. собственно наследственные болезни (например, фенилкетонурия, гемофилии);
болезни вследствие мутаций в соматических клетках - «соматические» (например, опухоли, некоторые болезни иммунной аутоагрессии); эти заболевания не передаются по наследству;
болезни вследствие комбинации мутаций в половых и соматических клетках (например, семейная ретинобластома).
Отдельно рассматривают летальные , сублетальные и гипогенитальные заболевания:
летальные болезни – приводят к гибели во время внутриутробного развития (например, моносомии по аутосомам, гаплоидии, большинство полиплоидии);
сублетальные заболевания – приводят к гибели индивида до периода полового созревания (например, наследственные иммунодефициты типа агаммаглобулинемии швейцарского типа, синдром Луи-Бар , некоторые гемофилии);
гипогенитальные заболевания – сочетаются с бесплодием (например, синдромы Шерешевского-Тернера, Кляйнфельтера ).
Деление наследственных болезней на генные и хромосомные неформальное: генные мутации передаются из поколения в поколение в соответствии с законами Менделя, в то время как хромосомные болезни , обусловленные анэуплоидиями, вообще не наследуются (летальный эффект с генетической точки зрения), а структурные перестройки (инверсии, транслокации) передаются с дополнительными перекомбинациями.
В основу генетической классификации наследственной патологии положен этиологический принцип - тип мутации и характер взаимодействия наследственных факторов со средой .
Генетические болезни соматических клеток выделены в самостоятельную группу после обнаружения в клетках при злокачественных новообразованиях специфических хромосомных перестроек, вызывающих активацию онкогенов. Эти изменения в генетическом материале клеток являются этиопатогенетическими для злокачественного роста и поэтому могут быть отнесены к категории генетической патологии .
Болезни, возникающие при несовместимости матери и плода по антигенам , развиваются в результате иммунной реакции матери на антигены плода. В целом эта группа составляет значительную часть патологии и довольно часто встречается в медицинской практике. Наиболее типичное и хорошо изученное заболевание этой группы - гемолитическая болезнь новорожденных , возникающая в результате несовместимости матери и плода по резус-антигену. Болезнь возникает в тех случаях, когда мать имеет резус-отрицательную группу крови, а плод унаследовал резус-положительный аллель от отца.
Клиническая классификация наследственных болезней ничем не отличается от классификации ненаследственных болезней по органному, системному признаку или по типу поражения обмена веществ .
Поскольку наследственные болезни едины по этиологическому принципу (мутации), основу их классификации составляет, прежде всего, системный и органный принцип : нервные; нервно-мышечные; кожные; глазные; опорно-двигательного аппарата; эндокринные; крови; легких; сердечно-сосудистой системы; мочеполовой системы; желудочно-кишечного тракта. Такой подход неоднозначен. Можно найти очень немного наследственных болезней, при которых избирательно поражается одна система. Большинство генных мутаций, а тем более хромосомные и геномные, вызывают генерализованное повреждение какой-либо ткани или захватывают несколько органов. Поэтому многие наследственные болезни проявляются в виде синдромов или комплекса патологических признаков.
Несмотря на очевидную условность, клиническая классификация помогает врачу соответствующего профиля концентрировать внимание на наследственных болезнях, встречающихся в практике данной специальности.
Патогенетическая классификация основана на выделении основного патогенетического звена.
Биохимическая классификация наследственных болезней, проявляющихся в нарушении обмена веществ, объединяет генетически й и клинический подход и проводится по типу повреждения первичного звена обмена .
В зависимости от характера нарушения обмена веществ различают:
· Наследственные дефекты обмена углеводов.
Галактоземия, нарушение метаболизма лактозы, мукополисахаридозы, нарушение расщепления полисахаридов.
· Дефекты обмена липидов и липопротеидов.
Сфинголипидозы - нарушения расщепления структурных липидов и другие формы нарушения обмена липидов .
· Дефекты обмена аминокислот.
Фенилкетонурия, альбинизм - нарушение синтеза пигмента меланина из тирозина и др.
· Дефекты обмена витаминов.
Гомоцистинурия - развивается как результат генетического дефекта кофермента витаминов группы В.
· Дефекты обмена пуриновых и пиримидиновых оснований.
· Дефекты биосинтеза гормонов.
Адреногенитальный синдром, тестикулярная феминизация.
· Наследственные дефекты ферментов эритроцитов.
· Коллагеновые болезни – дефекты биосинтеза и распада коллагена - структурного компонента соединительной ткани.
Болезнь Эллерса-Данло, болезнь Марфана и ряд других заболеваний.
Болезни, вызванные дефектами генов, кодирующих ферменты, наследуются по аутосомно-рецессивному типу , а вызванные генами, кодирующими модуляторные белковые функции или рецепторы, - по аутосомно-доминантному или аутосомно-рецессивному. Болезни, вызванные генами транскрипционных факторов, относятся к группе аутосомно-доминантных .В геноме человека данное правило иногда нарушается, что обусловлено различными мутациями в одном и том же гене. Это необходимо учитывать при медико-генетическом консультировании.
В основе болезней, патогенез которых приводит к аномалиям морфогенеза, лежит нарушение дифференцировки клеток, приводящее к врожденным порокам развития (полидактилия, синдромы Хол-Орама, Крузона и др.).
Однако подавляющее большинство моногенных наследственных болезней относится к третьей группе - обусловленных комбинацией нарушения обмена веществ и аномалий морфогенеза (муковисцидозы, ахондроплазии, мышечные дистрофии и др.).
Формирование наследственных болезней во времени также имеет закономерности, связанные с функцией первичного продукта гена:
болезни транскрипционных факторов развиваются внутриутробно;
патология ферментов - в течение первого года жизни;
рецепторов - в возрасте от 1 года до пубертатного периода;
модулярной белковой функции - в период до 50 лет.
ВИДЫ И ЗАКОНОМЕРНОСТИ ТЕЧЕНИЯ ГЕННЫХ БОЛЕЗНЕЙ
В широко известном научному и медицинскому сообществу и ежедневно пополняемом Каталоге наследственных заболеваний человека
(OMIM, Online Mendelian Inheritance in Man
Генные болезни - разнообразная по клиническим проявлениям группа наследственных заболеваний, обусловленных мутациями на генном уровне. Основой для объединения их в одну группу служат этиологическая генетическая характеристика и закономерности наследования в семьях и популяциях. Согласно многочисленным исследованиям разных наследственных болезней и генома человека в целом, можно говорить о многообразии видов мутаций в одном и том же гене. Любой из этих видов мутаций может вести к наследственным болезням. Даже одна и та же генная болезнь может быть обусловлена разными мутациями.
При изучении белковых продуктов мутантных генов выделяют две группы мутаций. Первая группа связана с качественными изменениями белковых молекул, т.е. наличием у больных аномальных белков (например, аномальные гемоглобины), что обусловлено мутациями структурных генов. Другая группа заболеваний характеризуется количественными изменениями содержания нормального белка в клетке (повышение или снижение), что обусловлено, чаще всего, мутациями функциональных генов, т.е. связано с нарушениями регуляции работы генов.
Мутации, вызывающие наследственные болезни, могут затрагивать структурные, транспортные, эмбриональные белки, ферменты. Наследственные аномалии могут возникать на всех уровнях регуляции синтеза белков , обусловленных соответствующими ферментативными реакциями:
претранскрипционном (осуществляется путем увеличения или уменьшения числа копий гена);
траскрипционном (генетические дефекты в спейсерах, интронах, транспозонах, регуляторных белках могут приводить к нарушению транскрипции всего гена, обусловливающие изменение объема синтеза соответствующего белка);
процессинга и сплайсинга про-и-РНК (нарушения на уровне разрушения неинформативных участков про-и-РНК и сплавления информативных участков);
трансляционном (нарушения на уровне непосредственной сборки белковой молекулы в рибосоме);
посттрансляционном (нарушения на уровне образования вторичной, третичной и четвертичной структуры белковой молекулы).
Поскольку мутации в индивидуальных генах являются этиологическим фактором генных болезней, то закономерности их наследования соответствуют менделевским правилам расщепления в потомстве.
При рассмотрении генных болезней как менделирующих признаков организма считается, что речь идет о так называемых полных формах , то есть формах, обусловленных гаметическими мутациями. Это могут быть новые или унаследованные от предыдущих поколений мутации. Следовательно, в этих случаях патологические гены присутствуют во всех клетках организма.
Однако теоретически можно представить появление и мозаичных форм . Любые мутации, в том числе генные, могут возникать на ранних стадиях дробления зиготы в одной из клеток, и тогда индивид будет мозаичен по данному гену. В одних клетках у него будет функционировать нормальный аллель, в других - мутантный или патологический.
В случае генных болезней клиническая картина заболевания может сформироваться вследствие патогенетических эффектов мутаций разных генов, то есть сходное фенотипическое проявление болезни может быть обусловлено несколькими различными мутациями. Следовательно, в одну группу попадут разные с генетической точки зрения случаи (мутации в разных локусах или разные мутации в одном локусе) – генокопии . Вместе с этим, хотя и редко, могут встречаться фенокопии генных болезней - случаи, при которых повреждающие внешние факторы, действующие, как правило, внутриутробно, вызывают болезнь, по клинической картине в общих чертах сходную с наследственной. Противоположное состояние, когда при мутантном генотипе в результате средовых воздействий (лекарства, диета и т. д.) болезнь не развивается, называют нормокопированием.
Понятия о гено- и фенокопиях дают врачу следующие возможности: поставить правильный диагноз; более точно определить прогноз состояния здоровья больного или вероятность рождения больного ребенка; в конкретном случае предупредить развитие болезни у ребенка, унаследовавшего патологический ген.
В соответствии с генетическим принципом классификации генные болезни делят на группы по типам наследования
· аутосомно-доминантные;
· аутосомно-рецессивные;
· Х-сцепленные доминантные;
· Х-сцепленные рецессивные;
· Y-сцепленные (голандрические) и митохондриальные.
Отнесение болезни к той или иной группе позволяет врачу сориентироваться относительно ситуации в семье и определить вид медико-генетической помощи.
В пятидесятых годах XX века ученые столкнулись со странным феноменом. Они обратили внимание на то, что некоторые вирусы по-разному заражают разные штаммы одной и той же бактерии. Некоторые штаммы - например, кишечной палочки - заражались легко и быстро распространяли инфекцию по колонии. Другие заражались очень медленно или вовсе были устойчивы к вирусам. Но однажды приспособившись к тому или иному штамму, в дальнейшем вирус заражал его уже без затруднений.
Биологам потребовалось два десятилетия, чтобы разобраться в такой избирательной устойчивости бактерий. Как выяснилось, способность определенных штаммов бактерий противостоять вирусам - ее назвали рестрикцией (то есть, «ограничением») - объясняется наличием у них специальных ферментов, физически разрезающих вирусную ДНК.
Особенность этих белков - ферментов рестрикции - в том, что они распознают небольшую и строго определенную последовательность ДНК. Бактерии «нацеливают» ферменты рестрикции на редкие последовательности, которых сами в своих генах избегают - но которые могут присутствовать в вирусной ДНК. Разные ферменты рестрикции опознают разные последовательности.
Каждый штамм бактерии имеет определенный арсенал таких ферментов и, таким образом, реагирует на определенный набор «слов» в геноме вируса. Если представить, что геном вируса - это фраза «мама мыла раму», то вирус не сможет заразить бактерию, опознающую слово «мама», но бактерия, нацеленная на слово «дядя», окажется беззащитной. Если же вирус сумеет мутировать и превратиться, скажем, в «баба мыла раму», то и первая бактерия потеряет свою защиту.
Почему открытие «бактериального иммунитета» оказалось на самом верху списка важнейших достижений молекулярной биологии? Дело не в самих бактериях и даже не в вирусах.
Отмерить кусочек ДНК
Ученые, описавшие этот механизм, почти сразу же обратили внимание на важнейшую деталь этого процесса. Ферменты рестрикции (точнее, один из типов этих ферментов) способны разрезать ДНК в четко определенной точке. Возвращаясь к нашей аналогии, фермент, нацеленный на слово «мама» в ДНК, связывается с этим словом и разрезает его, например, между третьей и четвертой буквой.
Таким образом, исследователи впервые получили возможность «вырезать» из геномов нужные им фрагменты ДНК. С помощью специальных «склеивающих» ферментов полученные фрагменты можно было сшивать - тоже в определенном порядке. С открытием ферментов рестрикции в руках ученых оказался весь необходимый инструментарий для «монтажа» ДНК. Со временем для обозначения этого процесса прижилась несколько другая метафора - генная инженерия.
Хотя сегодня существуют и другие методы работы с ДНК, подавляющее большинство биологических исследований последних двадцати-тридцати лет были бы невозможны без ферментов рестрикции. От трансгенных растений до генной терапии, от рекомбинантного инсулина до индуцированных стволовых клеток - любые работы, включающие генетические манипуляции, используют это «бактериальное оружие».
Знать врага в лицо
Иммунная система млекопитающих - в том числе человека - обладает как врожденными, так и приобретенными механизмами защиты. Врожденные компоненты иммунитета обычно реагируют на что-то общее, объединяющее сразу многих врагов организма. Например, врожденный иммунитет может распознавать компоненты клеточной стенки бактерий, одинаковые для тысяч разнообразных микробов.
Приобретенный же иммунитет полагается на явление иммунологической памяти. Он распознает конкретные компоненты конкретных патогенов, «запоминая» их на будущее. На этом основана вакцинация: иммунная система «тренируется» на убитом вирусе или бактерии, и в дальнейшем, при попадании в организм живого патогена, «узнает» его и уничтожает на месте.
Врожденный иммунитет - это пограничный пункт досмотра. Он защищает от всего сразу и при этом ни от чего конкретного. Приобретенный иммунитет - это снайпер, знающий врага в лицо. Как выяснилось в 2012-м году, нечто похожее есть и у бактерий.
![]()
Если рестрикция - это бактериальный аналог врожденного иммунитета, то роль приобретенного иммунитета у бактерий выполняет система с довольно громоздким названием CRISPR/Cas9, или «Криспер».
Суть работы «Криспера» заключается в следующем. Когда бактерия попадает под вирусную атаку, она копирует часть ДНК вируса в специальное место в собственном геноме (это «хранилище» информации о вирусах и называется CRISPR). На основе этих сохраненных «фотороботов» вируса бактерия затем изготавливает РНК-зонд, способный распознавать вирусные гены и связываться с ними, если вирус попытается снова заразить бактерию.
РНК-зонд сам по себе безобиден для вируса, но здесь в дело вступает еще один игрок: белок Cas9. Он представляет собой «ножницы», ответственные за разрушение вирусных генов - наподобие фермента рестрикции. Cas9 ухватывается за РНК-зонд и как бы на поводке доставляется к вирусной ДНК, после чего ему дается сигнал: резать здесь!
Итого, вся система состоит из трех бактериальных компонентов:
1) ДНК-хранилище «фотороботов» старых вирусов;
2) РНК-зонд, сделанный на основе этих «фотороботов» и способный опознать по ним вирус;
3) белковые «ножницы», привязанные к РНК-зонду и разрезающие вирусную ДНК ровно в той точке, с которой «фоторобот» был снят в прошлый раз.
Практически мгновенно после открытия этого «бактериального иммунитета» о бактериях и их вирусах все забыли. Научная литература взорвалась восторженными статьями о потенциале системы CRISPR/Cas9 как инструмента для генной инженерии и медицины будущего.
Как и в случае с ферментами рестрикции, система «Криспер» способна разрезать ДНК в строго определенной точке. Но по сравнению с «ножницами», открытыми в семидесятых, она обладает огромными преимуществами.
Ферменты рестрикции используются биологами для «монтажа» ДНК исключительно в пробирке: нужно сначала изготовить нужный фрагмент (например, модифицированный ген), и уже потом вводить его в клетку или организм. «Криспером» можно резать ДНК на месте, прямо в живой клетке. Это позволяет не просто изготавливать искусственно вводимые гены, но и «редактировать» целые геномы: например, удалять одни гены и вставлять вместо них новые. Совсем недавно о таком можно было только мечтать.
Как стало понятно за последний год, система CRISPR неприхотлива и может работать в любой клетке: не только бактериальной, но и мышиной или человеческой. «Установить» ее в нужную клетку довольно просто. Принципиально это можно делать даже на уровне целых тканей и организмов. В будущем это позволит целиком удалять из генома взрослого человека дефектные гены - например, вызывающие рак.
Допустим, присутствующая у вас в геноме фраза «мама мыла раму» вызывает в вас болезненную тягу к гендерным стереотипам. Чтобы избавиться от этой проблемы, вам нужен белок Cas9 - всегда один и тот же - и пара РНК-зондов, нацеленных на слова «мама» и «раму». Эти зонды могут быть любыми - современные методы позволяют синтезировать их за несколько часов. Ограничений по количеству вообще нет: «резать» геном можно хоть в тысяче точек одновременно.
![]()
Прицельная настройка организма
Но ценность «Криспера» не ограничивается «ножничной» функцией. Как отмечают многие авторы, эта система - первый известный нам инструмент, с помощью которого можно организовать «встречу» определенного белка, определенной РНК и определенной ДНК одновременно. Это само по себе открывает огромные возможности для науки и медицины.
Например, у белка Cas9 можно отключить «ножничную» функцию, а вместо этого привязать к нему другой белок - скажем, активатор гена. С помощью подходящего РНК-зонда получившуюся пару можно отправлять в нужную точку генома: например, к плохо работающему гену инсулина у некоторых диабетиков. Организуя таким образом встречу активирующего белка и выключенного гена, можно прицельно и тонко настраивать работу организма.
Привязывать можно не только активаторы, а вообще все что угодно - скажем, белок, способный заменить дефектный ген на его «резервную копию» с другой хромосомы. Таким образом в перспективе можно будет вылечить, например, болезнь Хантингтона. Главное достоинство системы CRISPR в данном случае - именно ее способность «отправлять экспедиции» к любой точке ДНК, которую мы можем запрограммировать без особых затруднений. В чем состоит задача каждой конкретной экспедиции - определяется только фантазией исследователей.
Сегодня сложно сказать, какие именно проблемы сумеет решить система CRISPR/Cas9 через несколько десятилетий. Мировое сообщество генетиков сейчас напоминает ребенка, которого пустили в огромный зал, до отказа забитый игрушками. Ведущий научный журнал Science недавно выпустил обзор последних достижений области под названием «The CRISPR Craze» - «Криспер-безумие». И все-таки уже сейчас очевидно: бактерии и фундаментальная наука в очередной раз подарили нам технологию, которая изменит мир.
В январе появились сообщения о рождении первых приматов, чей геном был успешно модифицирован системой CRISPR/Cas9. Мартышкам в качестве пробного эксперимента ввели мутации в два гена: один связанный с работой иммунной системы, а другой - ответственный за отложение жира, что непрозрачно намекает на возможное применение метода к homo sapiens. Возможно, решение проблемы ожирения методом генной инженерии - не такое уж далекое будущее.
Рекомендуем также
 Школьная форма в Америке и что такое dress code Школьная форма во франции
Школьная форма в Америке и что такое dress code Школьная форма во франции
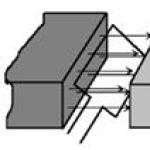 Эдс индукции в движущемся проводнике
Эдс индукции в движущемся проводнике
 Тема: Сюжетная аппликация
Тема: Сюжетная аппликация
 Конспект НОД по аппликации «Космическое путешествие Аппликация космическая ракета в подготовительной группе
Конспект НОД по аппликации «Космическое путешествие Аппликация космическая ракета в подготовительной группе
 Сонник рыба прыгает в руки
Сонник рыба прыгает в руки
 Сонник целоваться с парнем который нравится
Сонник целоваться с парнем который нравится